Contact us here!
Contact us for questions, general inquiries, or to request a quote.
By submitting this form, I consent to the processing of my personal data as explained in Ossiform's Privacy Policy.
Contact us for questions, general inquiries, or to request a quote.
By submitting this form, I consent to the processing of my personal data as explained in Ossiform's Privacy Policy.


Traditionally, microorganisms were studied as free-floating (planktonic) cells grown in cell culture plates, where they were described based on their growth characteristics in a nutritionally rich cell culture medium. While this setup has yielded invaluable insights into the microscopic world of bacteria, it only captures a glimpse of how these microbes really live, survive, and behave in nature and in the human body. In most environments, ranging from soil and aquatic environments to biological tissue and medical devices, microorganisms predominantly exist in their sessile lifestyle as biofilms (Donlan, 2002; Bjarnsholt, 2013). Biofilms are structured, surface-associated communities of one or more living microorganisms such as bacteria, fungi, and sometimes viruses in a self-produced extracellular matrix (ECM), also referred to as extracellular polymeric substance (EPS), designed to promote survival and persistence of the organisms. However, biofilms are not simply a cluster of cells and microbes, but a highly organized and dynamic system where microbial behavior and gene expression differ significantly from their planktonic counterpart. The matrix is typically composed of polysaccharides, proteins, lipids, and extracellular DNA, which provides structural integrity and mediate interactions within the community and the environment (López et al., 2010; Karatan & Watnick, 2009; Tolker‑Nielsen, 2015).
The ability of microbes to form biofilms is ancient and has been detected as far back as 3.4 billion years ago. Today, biofilm formation is recognized as a fundamental mode of microbial life, involving a regulated developmental process in which cells transition from planktonic growth to structured, surface‑associated communities (Monds & O’Toole, 2009; Noffke et al., 2013).
This mode of growth was already discovered in 1684, when scientist Antoine van Leeowenhook, using a simple microscope, observed dense microbial communities on the surface of animal teeth. He reported that the bacteria within the plaques of the teeth were more resistant to vinegar than the bacteria outside the plaques. This was an early observation of what is now recognized as a hallmark of biofilms: enhanced tolerance to antimicrobial agents (Donlan, 2002; Bjarnsholt, 2013).
Today, this tolerance is recognized as a major contributor to persistent and chronic infections, and it reflects biofilm‑specific features that collectively limit the effectiveness of antimicrobial treatments. With the increasing global threat of antibiotic resistance, understanding disease-causing microorganisms in their native biofilm state has becomes progressively relevant for both healthcare and the pharmaceutical research (Tolker‑Nielsen, 2015; Monds & O’Toole, 2009; Noffke et al., 2013). Understanding how microorganisms establish, maintain, and function within biofilms thus represents a key step toward deciphering microbial behavior in real‑world settings.
The development of biofilms is a complex and dynamic process that is commonly described as progressing through five stages:
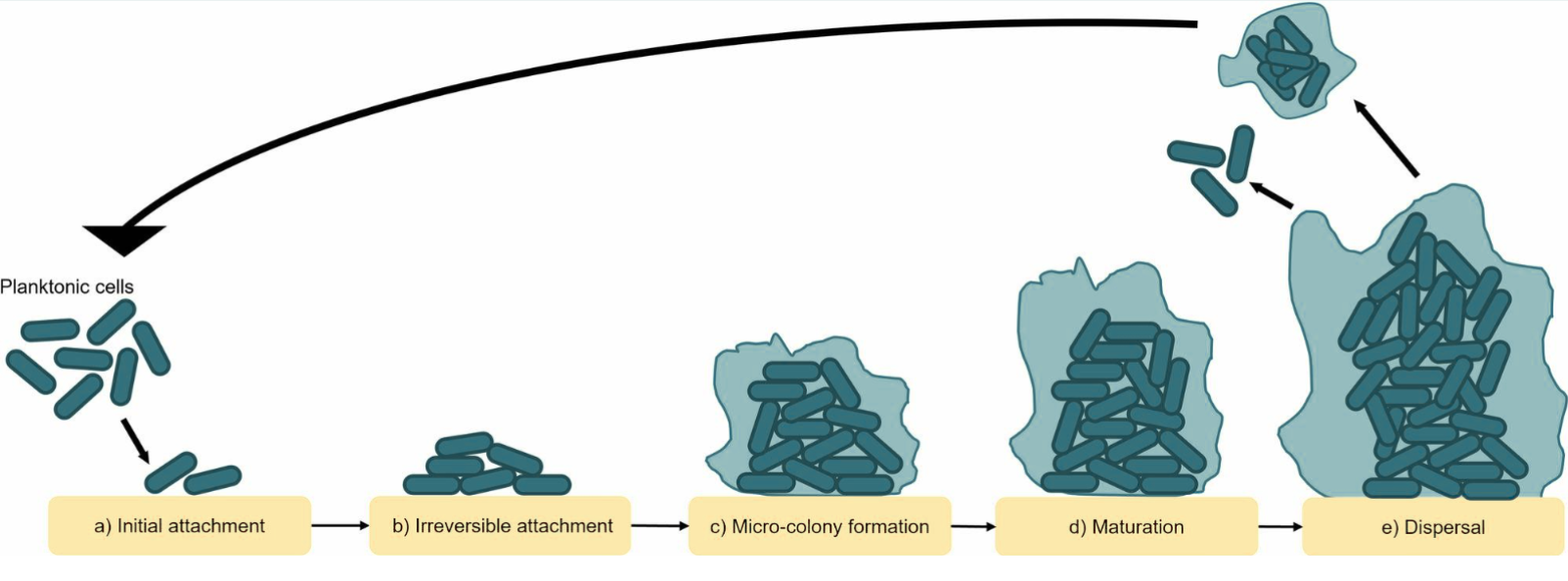
Figure 1: Mechanisms of biofilm formation. Adapted from Liu HY, Prentice EL, Webber MA. Mechanisms of antimicrobial resistance in biofilms. NPJ Antimicrobial Resistance. 2024;2:27, under the terms of the Creative Commons Attribution 4.0 International License (CC BY 4.0).
The first step of biofilm formation is the surface attachment. During the initial, reversible attachment stage, the microorganisms are loosely and reversibly associated with the surface, often in a polar manner. This is followed by irreversible attachment, during which cells loose polarity, adopt a flatter orientation on the surface, and commit to a sessile lifestyle. This stage is associated with internal cellular signaling that limits motility but increases biofilm matrix production as well as the development of increased tolerance towards external factors that may hinder biofilm formation. This internal signaling is triggered and amplified by each attachment/detachment event of the microbe, which subsequently initiates the early stages of matrix formation (Karygianni et al., 2020; Flemming et al., 2007).
Following successful attachment, the microorganisms proliferate and aggregate within the self-produced EPS, forming microcolonies. This early development phase depends on surface interactions and intercellular communication. As the biofilm matures, the EPS becomes increasingly important as it stabilizes the three-dimensional structure, retains signaling molecules for quorum sensing, and protects the embedded microbes from environmental stresses such as the immune system, antimicrobial agents, and oxidative damage. Within mature biofilms, microorganisms are spatially organized according to oxygen availability, nutrient gradients, metabolic activity, and waste accumulation (Karygianni et al., 2020; Flemming et al., 2007).
The final stage of the biofilm lifecycle is dispersion, during which the microbes detach from the biofilm and re-enter the planktonic phase. From here, a new biofilm formation cycle may begin. Dispersion can occur through active mechanisms, such as enzymatic degradation of the EPS, or passively in response to physical forces. Dispersion may be caused by population overgrowth, increased competition, nutrient limitations, metabolite accumulation, environmental changes or regulatory shifts that enhance motility and matrix breakdown (Karygianni et al., 2020; Flemming et al., 2007).
Biofilm development reflects a continuous and adaptive process as these stages are not rigid or irreversible but can overlap and vary depending on environmental conditions and microbial species. The ability to attach, grow, reorganize, and disperse allows microbial communities to persist across diverse and changing environments. Importantly, the same features that enable biofilm formation and stability also set the stage for the remarkable resilience observed in biofilm‑associated infections.
An important feature of biofilms is that biofilm-associated cells have properties that are distinct from their planktonic counterparts, including increased tolerance or even resistance towards antimicrobial agents. Diverse strategies for this resistance are employed, some being part of the fundamental properties of the biofilm resulting in intrinsic tolerance while others are active facilitators of the evolution of antibiotic resistance within and between species (Mah & O’Toole, 2001; Liu et al., 2024).
The biofilm matrix itself is a robust and protective barrier for the microorganisms embedded within. Firstly, it physically makes it more difficult for the antimicrobial agent to penetrate and reach the microbes as it slows the diffusion of the antibiotic agent. However, studies have shown that the biofilms not only slow the absorption of the antimicrobial agent but can actively reduce effective antimicrobial concentrations. Some antimicrobial agents form complexes with components of the matrix or are broken down by enzymes present in the biofilm, consequently reducing the concentration of the antibiotic that reach the target microbes (Mah & O’Toole, 2001; Liu et al., 2024).
Another mechanism of antibiotic resistance observed in biofilms is the inherent nature of the biofilm matrix and the growth of the microbes in the biofilm. Growing in a biofilm can lead to a general stress response and the microorganisms will almost inevitably experience some sort of nutrient limitation. When bacterial cells become stressed or starved, they forfeit propagation and their growth is slowed, which in turn is associated with increased antimicrobial resistance. Since many antimicrobial agents specifically target cellular processes associated with growth, when these are limited, the effects of the agent decrease. The slow-growing cells can even become metabolically inactive or dormant, and these are often found in the deeper layers of the biofilm. The inactive cells, referred to as “persister cells”, undergo phenotypic changes that allows them to survive with limited metabolic activity and ability to temporarily tolerate otherwise fatal concentrations of antibiotics. The existence of persister cells contributes considerably to the chronic nature of biofilm infections. After withdrawal of the antibiotic treatment, they can re-activate and repopulate the site of infection, causing further damage (Mah & O’Toole, 2001; Liu et al., 2024).
Another major contributor to antibiotic resistance in microbial communities is horizontal gene transfer. Horizontal gene transfer allows genes to be transferred between different species of bacteria making it possible for non-pathogenic and non-resistant microbes to obtain genes that make them resistant to antibiotics or gives them other advantages. Three primary mechanisms of horizontal gene transfer are described and include 1) transformation, where DNA from the surroundings are taken up by a bacteria, 2) transduction, where a bacteriophage facilitates the transfer of the genetic material from one bacteria to another, and 3) conjugation, where genes are transferred directly from one bacteria to another through direct contact between the donor and the recipient cell. All three mechanisms are relevant in biofilms, but conjugation is regarded as the most important for the transfer of antimicrobial resistance genes. Although horizontal gene transfer also takes place between planktonic cells, it occurs at higher rates in biofilms. Furthermore, biofilms can act as reservoirs for antimicrobial resistance genes, with surviving subpopulations contributing to their persistence, which further strengthens the antimicrobial resistance effects of the biofilm (Mah & O’Toole, 2001).
Taken together, the reduced susceptibility of biofilms to antimicrobial agents appears to be cause the interplay of multiple, overlapping mechanisms rather than a single dominant factor. This collective resilience enables biofilm‑associated microorganisms to persist despite antimicrobial intervention, particularly in environments where surfaces provide long‑term niches for microbial colonization.
In healthcare environments, the presence of biofilm‑forming microorganisms poses a particular risk due to the high density of vulnerable patients and the frequent use of antimicrobial agents. Hospitals and other healthcare facilities can act as reservoirs for microorganisms that are both biofilm‑forming and antimicrobial‑resistant, increasing the likelihood of persistent infections and transmission within clinical settings. Once introduced, such microorganisms may colonize surfaces, equipment, or patients, where biofilm formation enables long‑term survival despite routine cleaning and antimicrobial exposure (Donlan, 2002; Bjarnsholt, 2013).
Medical devices are especially susceptible to biofilm colonization because their surfaces provide stable interfaces that are rapidly conditioned by host proteins upon contact with bodily fluids. Even when devices are inserted under strictly controlled conditions, the introduction of a small number of microorganisms can be sufficient to initiate biofilm formation. For this reason, maintaining sterility during implant and device‑related procedures is critical, as early contamination greatly increases the risk of subsequent biofilm‑associated infection (Bjarnsholt, 2013; Arciola et al., 2018).

When biofilms establish on medical devices, they can serve as persistent sources of infection that are difficult to eradicate. In many cases, antimicrobial therapy alone is insufficient to fully clear device‑associated biofilms, allowing infections to persist or recur (Donlan, 2002; Bjarnsholt, 2013). As a result, effective treatment often requires invasive intervention as the most effective treatment of a biofilm infection is mechanical removal of the biofilm, including partial or complete removal of the affected device. Such procedures are associated with increased patient morbidity, prolonged hospitalization, and elevated healthcare costs, underscoring the significant clinical burden imposed by biofilm formation in medical settings (Donlan, 2002; Bjarnsholt, 2013; Percival et al., 2015).
Together, these challenges highlight the importance of preventing biofilm establishment on medical devices through rigorous infection‑control practices, strict sterility during implantation, and continued efforts to understand how biofilms interact with artificial surfaces in clinical environments.
Due to the significant human health implications of biofilms, research concerned with biofilm formation, prevention, and treatment can potentially enhance medical and dental practices.
Since biofilm has been found to be part of many chronic infections, knowledge on how biofilm may contribute to the pathogenesis of disease is important for the development of effective treatments for biofilm associated infections.
The field of research mostly investigates the formation of biofilms on implants and solid support matrices, or scaffolds, to obtain knowledge on the communication between the biofilm cells. Namely, in the hope of finding a solution to the antibiotic resistances or to gain a better understanding of the complex community.
To reach an understanding of biofilm formation mechanisms, the P3D scaffold can be used to investigate microbe attachment and how biofilm-residing cells interact with 1) each other, 2) synthetic bone implants, and 3) bone cells. Moreover, the lifelike 3D environment can be used to test the effect of antimicrobial treatments.
If you have any questions, please email research@ossiform.com.